エンドファイト −その特性と利用−
はじめに
芝と言うのは、割合身近かな植物である。公園やグラウンド、河川敷、ゴルフ場などに広がる芝は、都会人にとってはイネよりも触れる機会の多い植物であろう。しかし、この植物に対する研究は、他の栽培植物に比べて大きく立ち遅れているのが実情である。明治以降の120年余、日本における農学は、イネを始めとする食用植物がその研究の中心であり、芝のように人間の口に入らない植物の研究者は極端に少なかった。
しかし、芝を取り巻く社会情勢は近年急激に変化している。日本が経済大国化すると共に、日本人の生活にも余暇を楽しむ”ゆとり”が増え、これが芝と接する機会を増加させた。ゴルフは今や大衆スポーツと化し、Jリーグは毎週末のスポーツ紙の一面を飾るようになった。こうした変化の影響で、今まで公園の片隅で小さくなっていた芝が一躍脚光を浴びるようになり、それと共に様々な問題点の改善もまた、社会の側から求められる様になった。現在”シバ”と言う植物に対する研究の進展が強く望まれているが、それはこのような非常に社会的な理由からである。
今回、「生物機能利用の最前線」と言うテーマの下に寒地型芝草のエンドファイトをご紹介する機会を得たが、テーマとの関連性の点でご批判を受ける事を承知の上で、芝を取り巻く諸問題を総合的に取り上げ、その中でエンドファイトが持つ可能性について考察してみたい。
芝生の効用
ここまで「芝」と言う言葉を無造作に用いてきたが、ここで言葉の定義をしておきたい。一般に「芝」と言う言葉は、芝生ならびにそこに生えている植物を指す、ややあやふやな語である。以後、単一または少数の草本によって被覆された土地を「芝生」、芝生を構成している草種を「芝草」と呼ぶ事にする。
さて、芝生の最大の効用は、芝草によって覆われた地面の上空に何もない空間を作り出す事にある(日本の主要な自然植生である森林の中でゴルフやサッカーをプレイするとどうなるかを想像していただければ、この何もない空間がどれほどの意味を持つか、ご理解いただけると思う)。もちろん、コンクリートや人工芝によっても同様の空間は確保できるが、そこで転倒したらどうなるか。緑色がもたらす心理的な安心感・解放感。直射日光下での表面温度などを総合すれば、芝生の有利性は明らかである。
「芝草」の分類
「芝草」、すなわちこうした芝生に植える植物としては、どのような植物が望ましいであろうか。公園のお弁当広場ならば、草丈の低い草本が密生していれば充分であろう。しかし、スポーツターフの様に、一定以上のクオリティと均一性を要求されるとなると話は違ってくる。永年性である事。密植に耐える事。地表面の被覆力が強い事。踏圧に強い事。再生力が強い事。葉が細く均一な事。葉色が良い事。などが芝草として持つべき要件である。結果として、芝草はほぼ全てがイネ科に属する。主要な芝草草種を
Table 1 に示した。このうち、スズメガヤ亜科とキビ亜科に属するものは暖地型芝草(warmseason
turfgrass)と呼ばれ、C4光合成を行う。ウシノケグサ亜科の草種は寒地型芝草(coolseason
turfgrass)と呼ばれ、C3光合成である。
Table 1 主要なイネ科芝草の系統分類表
芝草栽培と気候
ゴルフ発祥の地はスコットランドである。なぜスコットランドでゴルフが生まれたのであろうか。スコットランドの気候は冷涼小雨でかつ年格差が少ない。こういった気候下で放牧圧をかけると(スコットランドは牧畜が盛んだった)、植性は自然と草地になる。ゴルフの誕生には自然環境と言うバックボーンが存在したのである。全英オープンの舞台となるリンクスコースは、人間が造ったのではなく天与の物だったのである。
翻ってわが国はどうか。南北に長い国土には様々な気候が広がるが、総じて年格差が大きい(Fig.1)。これは、一つの草種で一年間緑を保つのが難しい事を示している。スコットランドならば寒地型芝草を植えておけば一年中緑(エバーグリーン)だし、シンガポールのように一年中暑ければ、暖地型芝草でエバーグリーンを達成できるのだが、そうも行かない。日本在来種であるシバ属(Zoysia)の芝草は、暑さや乾燥に強いため、わが国では最も広く使用されているが、Zoysiaには冬期に休眠して葉が枯死する欠点がある(ほふく茎や地下茎が越冬し、翌春に芽を出す)。日本の気候に上手く適応しているわけだが、利用する側にとってはあまり好ましい事ではない。寒地型芝草を用いれば冬期に緑色を維持することは可能だが、今度は高温多湿の日本の夏を越えさせるのが難しい。両者を混植する方法もあるが、ひとつ間違えると共倒れになる可能性もあるので、キメ細かい管理が可能な場所に限られる。
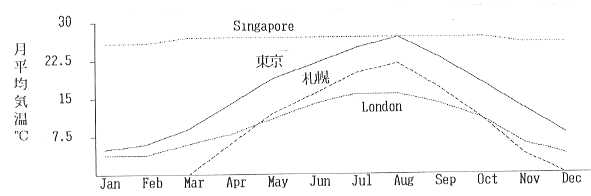
Fig.1 東京、札幌、London及びSingaporeの月平均気温の年間変動
そもそも夏期の気温が高く降水量の多いわが国では、芝生や草地を維持管理して行くのは難しい。北海道、東北や山地と言った寒冷地はともかく、関東以西の平野部で芝生を維持して行くのは、農業における適地適作の原則を大きく逸脱するものである。当然病虫害や雑草の発生が多くなる。と言って、ゴルフやサッカーを芝生以外の場所で行う訳にもいかない。芝草をめぐる諸問題の根底にはこの事があると言うのが、筆者の考えである。
芝草の総合防除
上述のように日本の芝草栽培(芝生の維持管理)は、その年の天候にもよるが、病虫雑草害との闘いになる。さらに、芝草の病原菌・害虫は土壌内に生活するものが多いため、発生予察が難しい。このため最近まで、特にゴルフ場などでは農薬を予防的にスケジュール散布する事によってこれらを制御する場合が多かった。しかしご承知の様に、ゴルフ場における農薬の使用は近年特に問題視されており、今後はかなり制限されてゆく方向にある。これは世界的傾向で、このため芝草においても「総合防除(Integrated
Pest Management; IPM)」が叫ばれている。これには、病虫害の発生予察、栽培方法の改善(これを唄い文句に、やたらと芝生用の土壌改良資材が現われたが)、そして耐病虫害品種の育種などが上げられる。
今回取り上げるエンドファイトと言う糸状菌(カビ)は、芝草の体内に共生し、種子を通じて次代に伝わる。そして、宿主に耐虫・耐病性や耐ストレス性を付与すると言う興味深い特徴が、芝草の品種改良に役立つのではないかと近年注目を集めている。以下に、このエンドファイトと言う風変わりなカビをご紹介したい。
エンドファイト研究の歴史
と言うわけで。やっとエンドファイト(endophyte)にたどり着いた。エンドファイトとは、「endo;内側を意味する接頭語+phyte;植物を意味する接尾語」を合わせたもので、広義には植物の内生共生微生物を指すが、芝草においてはAcremonium属の共生糸状菌を意味する場合が多い。本稿でも、エンドファイトと言った場合には、本属の共生糸状菌を指す事にする。
寒地型芝草の一部の草種に内部共生糸状菌が存在することは19世紀末に既に報告があり、この菌が種子伝染する事も1930年代に既に知られていた1)。その後は忘れ去られたまま時間が過ぎて行ったが、1977年になって、ペレニアルライグラス(PRG)やトールフェスク(TF)を飼料として与えたウシやヒツジに起こる中毒症状が、共生しているエンドファイトが生産するアルカロイドに起因するものである事が報告され2)、一躍注目を浴びるようになった(芝草と牧草は共通する草種が多い)。そして80年代に入ると、様々な面からエンドファイトに関する研究が進められ、この菌が持つ種々のユニークな特性が明らかにされてきた。
エンドファイトの生活史
エンドファイトと芝草の共生関係がいつ頃成立したかは不明だが、かなり長い時間が経過しているのは確かである。この長い共生の結果、エンドファイトは宿主植物外での生活能力を失ってしまい、宿主の体内でのみ生存が可能である。当然植物体への侵入能も失ってしまい、エンドファイト感染個体(endophyte
infected plant(s); EI)と非感染個体(endophyte free plant(s); EF)を混植しても、EFがエンドファイトに感染するような事はない。種子を通じて次世代に伝染する事によってのみ、エンドファイトはその種を維持している。
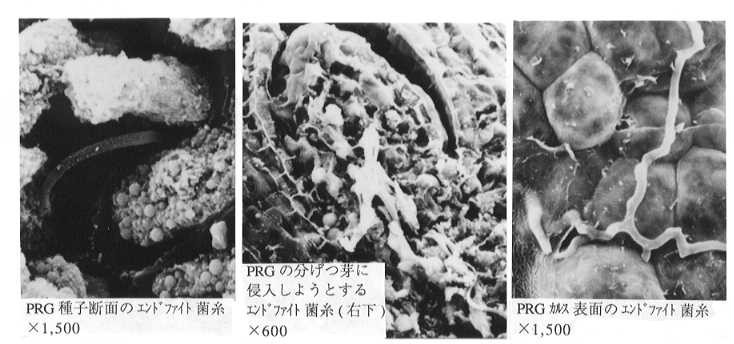
種子中のエンドファイト菌糸は、胚乳の細胞間隙に存在する。胚の中まで侵入することはないようである。種子が低温で保管される場合には、エンドファイトの感染率はほとんど低下しないが、室温で保管される場合には、2〜3年程度で死滅する。種子が水と温度を得て発芽すると同時に、エンドファイトも成長を開始する。植物体中では成長点付近に分布し、ここから展開して行く新葉や分げつに分配される。ただ、この場合でも細胞内や維管束内に侵入することはなく、細胞間隙を進展して行く。そして、出穂が起こると穂軸内を上昇し、種子に侵入する。こうして次世代に伝わると共に、
生活環を完成させる(Plate 1)。
Plate 1 植物体内のエンドファイト菌糸
エンドファイトの生育スピードは非常に遅い。培地上では 0.1-0.5mm/day
程度である。生活の場を宿主に依存しなければならないエンドファイトにとって、宿主の迷惑にならない程度に生育スピードを落とす必要があったのだろう。このためエンドファイトの分離には、通常4〜8週間を程度を要する。また、芝草種子から誘導した、見かけ上エンドファイトフリーのカルスを継代しているとエンドファイトが出てくる事があり、芝草組織培養上の問題点となっている。
エンドファイトの宿主植物
さて、このAcremonium属エンドファイトは、全ての芝草草種に共生しているかと言うと、そうではない。寒地型芝草のFestuceae連(Fig.
1)に限られており、ベントグラス(Agrostis)などにはAcremonium属エンドファイトの存在は確認されていない(最近見つかったと言う情報もあるが)。何故、この様な事が起こるのだろうか。エンドファイトは花粉に侵入できないため、種子伝染すると言っても常に母親から子へと伝わる。葉緑体や、ミトコンドリアと同様に母系遺伝である(葉緑体やミトコンドリアの起源に関する共生説を考え併せると面白い)。これは想像でしかないが、遠い過去のある時点で、エンドファイトと共生関係に入った芝草を母系とする子孫だけが、エンドファイトを有しているのであろう。
エンドファイトの効果
さて、Acremonium属エンドファイトが共生する事が、芝草やそれを利用する人間にとってどう言う意味を持つのであろうか。現在までに報告のあるエンドファイトの効果をTable
2にまとめたが、実に多岐に渡っている。
Table 2 エンドファイトが植物に及ぼす効果
家畜毒性、 耐虫性、 生育向上、 ストレス耐性、
耐線虫性、 耐病性、 耐酸性雨性
家畜に対する毒性については、エンドファイトが注目を集める発端になったものであり、最も研究が進んでいるが、芝として見た場合にはあまり問題にならないので、ここでは割愛する。なお、Fig.
2の1(TFから)、2-3(PRGから)などが毒性物質として同定されている。なお、この他にも多数の物質の報告がある。
耐虫性については、摂食性吸汁性を問わず多くの昆虫に効果があり、日本で大きな問題になっているシバツトガにも効果がある3)4)。これら耐虫性の主要因は摂食忌避であり3)4)、その原因物質についても同定が行われている(Fig.
2の4-6)。アブラムシでは、種5)や生育ステージ6)によって、感受性が異なることが知られている。また、家畜毒性と耐虫性の原因物質が異なる事から、害虫にのみ効果を示す菌株を持つ牧草品種の育成も行われている。
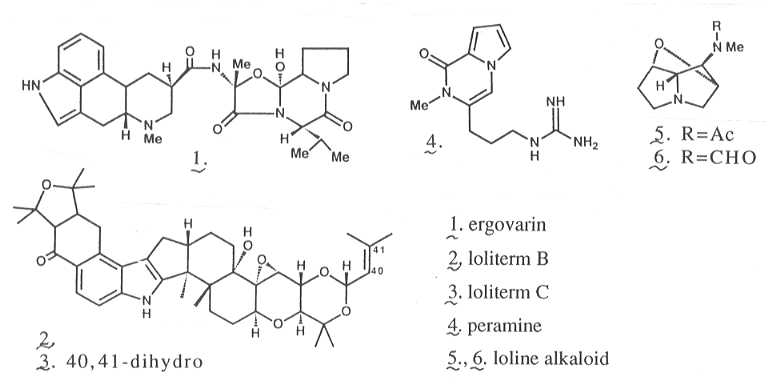
Fig. 2 エンドファイトが生産する生理活性物質
Arachevaletaらは7)、TFにおいてEIがEFよりも乾物生産量が最大67%増大し、これはグルタミンシンセターゼ活性の増大によるものとしている。また、乾燥条件下での生存率が向上し。これは葉の形態変化によるものであると考察している。一方で、EIとEFで乾燥抵抗性に差はないと言う報告もある8)。こうした環境抵抗性は微妙なものであり、試験条件によって差が出る事が多いので、慎重に検討すべき必要があるだろう。
日本においては芝草の線虫害はあまり問題になっていないが、米国ではからに大きな被害を及ぼしている(もっとも、日本には線虫の研究者が少ないため報告がないだけで、実際には被害が出ているのかも知れない)。EIを植えた芝生では、EFを植えた場合よりも被害が少なく、土壌中の植物寄生性線虫も少なかった9)。根におけるエンドファイト菌糸の量は、地上部に比べると著しく少ないが10)、にもかかわらずこのような効果を発揮するメカニズムはまだ分かっていない。
耐病性については、PRGのRhizoctonia zeae による苗立枯れが軽減された11)との報告がある一方で、TFのRhizoctonia
blight には効果がない12)との報告もあり、それほど強いものではないらしい。また、EI
はEF よりBarley Yellow Dwarf Virus の感染率が低いとの報告があり13)、メカニズムが注目される。
さらにモデル実験ではあるが、酸性の水を土壌に潅注した場合には、EI はEF
よりも耐性があった14)との報告がある。
エンドファイトの育種への利用
以上説明してきた様に、エンドファイトは芝生の管理者にとって有益であり、また種子伝染(実際上は遺伝と言っていい)すると言う特徴は、芝草の育種に利用可能である。事実、エンドファイトの研究が進められて行った80年代の後半から、エンドファイト含有を唄い文句にした品種が育成されるようになり、現在では新しく発売される芝用のPRGやTFのかなりの品種がエンドファイトに高率で感染しており、またこれをセールスポイントにしている。これらの品種は、育種の過程でエンドファイトの有無を光学顕微鏡でチェックし、感染個体を選抜する事により育成されてきた。
筆者の所属する(株)ジャパン・ターフグラス(JTG)では、バイオテクノロジーを用いた芝草の新品種の育成を研究目標の一つの柱に置いている。以下に、筆者が行ってきた、エンドファイトの芝草への導入についてご紹介する。
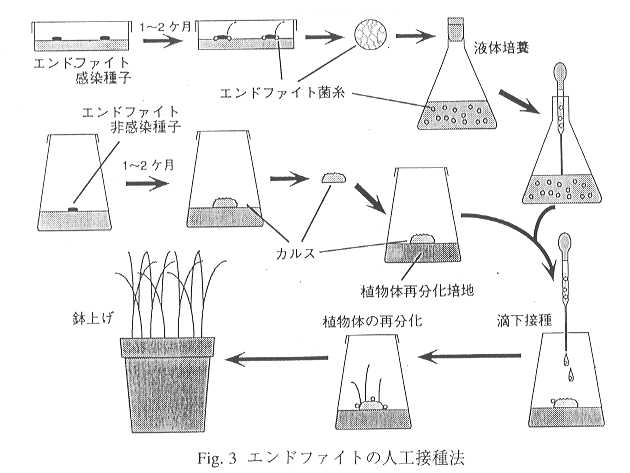
エンドファイトは植物への侵入能を喪失しているため、植物表面に菌糸を散布するような方法では感染しない。そこでエンドファイトの接種方法として、植物体に付傷接種する方法15)と、カルスに対して接種した後、植物体を再分化させる方法16)が考案された。育種を行うにあたっては、極力多くの個体の中から優良な個体を選抜する事が重要である。そこで、一度に多くの感染個体を得られるカルスに対する接種方法を選び、対象草種としてこの方法ではまだ成功していなかったPRGを選んだ。接種方法の概要をFig.
3に示した。方法自体は簡単であるが、エンドファイトの菌糸は生育が遅く、分離に2ヶ月程度を要すること、さらにカルスを誘導し、その中から再分化能の高い株を選抜するするのに6ヶ月程度かかるなど、かなり気の長い作業が必要である。
エンドファイトの接種を行うに当たって、さらにクリアしなければならない問題点がいくつかある。一つは、接種するカルスが真にエンドファイトフリーである事をチェックする方法。二つには、接種後再分化させた植物体中に糸状菌の菌糸が観察されたとして、それがAcremonium
属エンドファイトであると同定する方法である。見かけ上エンドファイトフリーと思われるカルスを継代していると、エンドファイトが出てくる例がある。このようなエンドファイトが潜在感染しているカルスを再分化させて、エンドファイトが入ったと言ったのではお話しにならない。また、芝草に共生する糸状菌は実はAcremonium
だけではない17)18)。再分化植物体中に菌糸が観察されたとしても、それがAcremonium
であると証明するためには再分離を行う必要があるが、これは時間のかかる作業である。これらの問題点を解決するため、血清学的方法を用いる事にした。
エンドファイト菌糸を液体窒素下で磨砕し、これを抗原としてウサギに免疫した。このウサギ血清はエンドファイトとのみ特異的に反応し19)、この血清を用いてELISAを行う事により、カルスのエンドファイト潜在感染の検定が可能となった(Table
3)。また、この血清を用いて蛍光抗体染色を行う事により、植物体中のエンドファイトを蛍光顕微鏡下で直接検出できるようになった(Plate
2)。これらの技術を組み合わせた結果、PRGのエンドファイト非感染品種であるManhattan
IIにエンドファイトを感染させる事ができた20)。
Table 3 ELISAによるカルスからのエンドファイト検出
|
カルス誘導品種
|
A405/600
|
|
Pinnacle
|
1.817, 1.240, 0.138, 0.098, 1.391,
0.138, 1.304, 0.163, 0.173, 0.148,
0.109, 0.128, 0.140, 1.418, 1.761, 0.155 |
|
Repell
|
0.081, 0.133, 0.102, 0.166, 0.107 |
|
Manhattan II
|
0.122, 0.137, 0.105, 0.177, 0.127,
0.126, 0.151, 0.106, 0.122 |
エンドファイト潜在感染カルスでは、A405/600>1 であった。

Plate 2 蛍光抗体染色で蛍光を発するエンドファイト菌糸
エンドファイトの非宿主への導入
エンドファイトの宿主範囲が限られている事は既に述べた。エンドファイトの主な宿主であるPRGやTFの日本における栽培適地は、北海道、東北及び中部地方の高地である。これより南になると、夏の高温多湿のため栽培が難しい。高度な管理を行えば栽培は可能であるが、栽培コストを考えると割に合わない場合が多い。ゴルフのグリーンに用いられるクリーピングベントグラスや、日本で最も広く栽培されているZoysia
属の芝草にエンドファイトを導入することができれば、エンドファイトの利用価値は大いに上がる。JTGにはこれらの草種のカルス−再分化系があるので、現在この系を用いて、クリーピングベントグラスならびにノシバへのエンドファイト接種を行っている。これらの草種はほふく茎を持っているので、栄養繁殖が可能である。このため次世代への移行を考えなくてもよい言う利点がある。
しかし、まだ実施例が少ないので結論は出せないが、本来の宿主以外への草種に感染させるのは、やはり難しいようである。今後は、これらの草種へのエンドファイトの導入を試みると共に、耐暑性の強いPRGやTFを作出し、これにエンドファイトを感染させる方法で、エンドファイトの利用を進めて行きたい。また、Acremonium
属エンドファイト内においても菌株による変異があることが報告されているので21)、優良な菌株の選抜についても行っていく予定である。
おわりに
植物に病気を引き起こす微生物、あるいは根粒菌のように外見上の変化を引き起こす微生物については、これまで多くの研究がなされてきた。しかし、広い意味でのエンドファイト−共生微生物−に関する研究は、近年までほとんど手付かずのままであった。が、Acremonium
属エンドファイトが発端となって、芝草/牧草の多くの草種で共生微生物の探索が行われ、かなりの草種に様々な種類の微生物が共生している事が判明してきた。そして、Acremonium
属エンドファイトについては、この菌の共生がもたらす多くのメリットが判明し、実用化されている。
では、芝草/牧草以外の植物には、共生微生物は存在しないのであろうか。これについては、これまで組織だった探索が行われていないため詳細は不明である。しかし、共生微生物の存在がイネ科植物に特異的と考えるよりは、多くの植物に普遍的に存在すると考える方がより自然であろう。種子伝染のレベルまで緊密化した共生と言うのは確かに珍しいかも知れないが、後天的な植物と微生物の共生関係と言うのは、意外にありふれた事のように筆者には思われる。今後、芝草に限らず栽培植物全般において、こうした観点からの生物防除にに関する研究が発展する事を期待している。
引用文献
1) Sampson (1937). Transactions British Mycological Soc.
23:316-.
2) Bacon et al. (1977). Appl. Environ. Microbiol. 34:106-.
3) 神田ら(1992). 日植病報 58:587.
4) Rowan and Gaynor (1986). J. Chem. Ecol. 12:647-.
5) Johnson et al. (1985). Appl. Environ. Microbiol. 49:568-.
6) Kindler et al. (1991). J. Econ. Entomol. 84:685-.
7) Arachevaleta et al. (1989). Agron. J. 81:83-.
8) White et al. (1993). Crop Sci. 32:1392-.
9) Kimmons et al. (1990). Plant Dis. 74:811-.
10) 笠原ら(1993). 日植病報 59:281.
11) Gwimm and Gavin (1992). Plant Dis. 76:911-.
12) Burpee and Bouton (1993). Plant Dis. 77:157-.
13) Mahmood et al. (1993). Plant Dis. 77:225-.
14) Latch and Christensen (1985). Ann. Appl. Biol. 107:17-.
15) Cheplick (1993). International J. Plant Sci. 154:134-.
16) Johnson et al. (1986). Planr Dis. 70:380-.
17) Philipson (1989). N.Z. J. Bot. 27:513-.
18) Pereria et al. (1993). Mycologia 85:362-.
19) 笠原ら(1992). 日植病報 58:102.
20) 笠原ら(1992). 日植病報 58:568.
21) Christensen et al. (1991). Mycological Res. 95:918-.